Article écrit à 6 mains, par Nicobola, Boris et Aurélide.
« Joyeux anniversaire ! »
 |
| Source |
Cette phrase, on l’entend au moins une fois par an. Et qui dit anniversaire dit… oui, gâteau, on sait Donald, tu as toujours faim. Oui, fiesta, on sait Nico, tu aimes faire la bringue !!! Mais surtout… oui Aurélide, tu as raison : les cadeaux !
Souvenez-vous, quand vous étiez petits, ouvrant de grands yeux devant l’amoncellement de présents… Dès lors que vous aviez repéré un emballage qui vous semblait prometteur, plusieurs choix s’offraient à vous. Certains palpaient le papier cadeau, tandis que d’autres secouaient les boites mystérieuses pour essayer d’en deviner le contenu… d’autres encore se précipitaient et arrachaient l’emballage d’un coup sec pour révéler ce qui y était dissimulé !
Mais que viennent faire les cadeaux d’anniversaire sur ce blog ? Eh bien, comme les enfants qui cherchent à deviner ce qui se cache sous les emballages, les naturalistes utilisent différentes techniques pour chercher à savoir ce qu’ils ont entre les mains en faisant intervenir l’ensemble des cinq sens.
Nous allons vous présenter ici comment identifier les êtres vivants qui nous entourent à l'aide de tous les moyens que nous avons à notre disposition, sans avoir besoin d'appareils technologiques perfectionnés.
La vue :
Commençons avec le sens que nous utilisons le plus en tant que primates : la vue.
La majorité des critères qu’on utilise pour distinguer les espèces entre elles sont des critères que l’on peut observer. C’est bien pour ça que tous les guides naturalistes utilisent largement des illustrations et des descriptions visuelles. Cependant les formes et les couleurs ne sont pas les seules caractéristiques visibles des êtres vivants : d’autres indices visuels auxquels on pense moins peuvent nous aider grandement à l’identification de certains animaux, comme le mouvement (déplacement, comportement) ou les traces qu’ils peuvent laisser dans leur environnement ! Voici quelques exemples :
Il existe dans les mares une diversité insoupçonnée car la plupart du temps quasiment invisible tant les animaux sont minuscules. On trouve dans ces mares trois taxons de crustacés majoritaires : les cladocères (dont font partie les daphnies), les copépodes et les ostracodes. Il est parfois difficile de différencier ces tous petits animaux à l’œil nu. Evidemment l'identification est facilitée par l’utilisation d’une loupe. Pourtant, plusieurs autres caractères peuvent permettre de les différencier sans avoir besoin de matériel optique, ni de devoir connaitre la morphologie très précise des espèces. Les cladocères nagent en battant des antennes par petits bonds verticaux tandis que les copépodes nagent horizontalement par à-coups successifs aussi grâce à leurs antennes. Quant aux ostracodes, ils ont plutôt tendance à rester au fond et nagent avec leurs pattes de manière continue. Voilà comment l’observation de leur mouvement peut aider à identifier des organismes.
Copépodes :
La majorité des critères qu’on utilise pour distinguer les espèces entre elles sont des critères que l’on peut observer. C’est bien pour ça que tous les guides naturalistes utilisent largement des illustrations et des descriptions visuelles. Cependant les formes et les couleurs ne sont pas les seules caractéristiques visibles des êtres vivants : d’autres indices visuels auxquels on pense moins peuvent nous aider grandement à l’identification de certains animaux, comme le mouvement (déplacement, comportement) ou les traces qu’ils peuvent laisser dans leur environnement ! Voici quelques exemples :
Il existe dans les mares une diversité insoupçonnée car la plupart du temps quasiment invisible tant les animaux sont minuscules. On trouve dans ces mares trois taxons de crustacés majoritaires : les cladocères (dont font partie les daphnies), les copépodes et les ostracodes. Il est parfois difficile de différencier ces tous petits animaux à l’œil nu. Evidemment l'identification est facilitée par l’utilisation d’une loupe. Pourtant, plusieurs autres caractères peuvent permettre de les différencier sans avoir besoin de matériel optique, ni de devoir connaitre la morphologie très précise des espèces. Les cladocères nagent en battant des antennes par petits bonds verticaux tandis que les copépodes nagent horizontalement par à-coups successifs aussi grâce à leurs antennes. Quant aux ostracodes, ils ont plutôt tendance à rester au fond et nagent avec leurs pattes de manière continue. Voilà comment l’observation de leur mouvement peut aider à identifier des organismes.
Copépodes :
Daphnies :
Ostracodes :
Anecdotes de Nicobola à propos de petites bêbêtes méconnues :
Un autre petit exemple - qui me tient à cœur car j’ai travaillé dessus - ce sont les Limnodriloidinae, qui font partie d'un taxon plus large dit des « clitellates ». Les clitellates contiennent entre autres les sangsues et les vers de terre. Rien de bien appétissant là dedans. On trouve en grand majorité dans ce groupe des petits vers à la limite du microscopique, très difficiles à différencier. En pratique on les colore et on observe au microscope leurs organes génitaux (oui les scientifiques sont des pervers). Plus particulièrement, ce sont des caractères tels que « l’entonnoir spermatique » ou la « vasa deferentia » que l’on observe. Cependant mon chef, lui, pouvait reconnaître les vers vivants (voir même morts) presque toujours au niveau du genre sous une loupe binoculaire ! Pour vous dire, les gens qui savent faire ça dans le monde doivent se compter sur les doigts d’une main. Son secret ? La forme générale certes mais aussi, le mouvement des individus lorsque ceux-ci sont encore vivants. Par exemple les Enchytraeidae (de petits vers blancs très communs dans le sol, comme le ver grindal pour les aquariophiles) ont une carapace plus rigide, leurs mouvements seront donc en général plus lents et moins souples. Les individus du genre Pristina, qu’on trouve souvent dans les mares, sont bien plus actifs et vont se promener un peu partout.
Voici un exemple, un ver du genre Pristina :
Un jour mon chef m’a envoyé en Afrique du Sud chercher (entre autres) ces fameux Limnodriloidinae. Mais comment reconnaître les Limnodriloidinae ? « Facile ! » me dit-il, « Ils sont rouges (comme beaucoup d’autres) et ils s’enroulent sur eux même et explorent les alentours avec leur tête. » Drôle de comportement ! Ca n’a pas raté ! J’en ai trouvé qui répondaient à cette description, que j’ai filmé, identifié comme ce groupe grâce à la vidéo, puis confirmé grâce à la morphologie de l’appareil génital plus tard et au final grâce à l’ADN aussi. Voici une vidéo pour vous convaincre (probablement la plus intéressante que vous n’ayez jamais vue !) :
C'est au tour d'Aurélide de prendre la parole !
Dans un tout autre contexte, on nous apprend en forêt à faire la différence entre des traces de différents animaux, par exemple entre un chevreuil ou un renard. Vous avez aussi peut-être appris à différencier leurs empreintes de pas sur le sol ou à détecter leurs crottes et autres déjections. Vous pouvez par ailleurs lire cet article de Taupo. Mais mis à part les mammifères craintifs qui sortent principalement la nuit, il existe une myriade d’animaux difficiles à trouver dont les traces sont parfois les seules choses qui trahissent leur présence.
Dans beaucoup d’environnements, on peut aussi utiliser le critère des traces pour identifier des animaux. Si vous allez régulièrement à la plage, vous avez sûrement dû voir des pêcheurs chercher des vers pour appâter leurs hameçons à l’aide d’une pelle ou d’une bêche : ils s'en vont chercher des vers marins fouisseurs. Ça n’a peut-être pas l’air, mais finalement, c’est très simple d'en trouver quand on sait ce qu’il faut regarder.
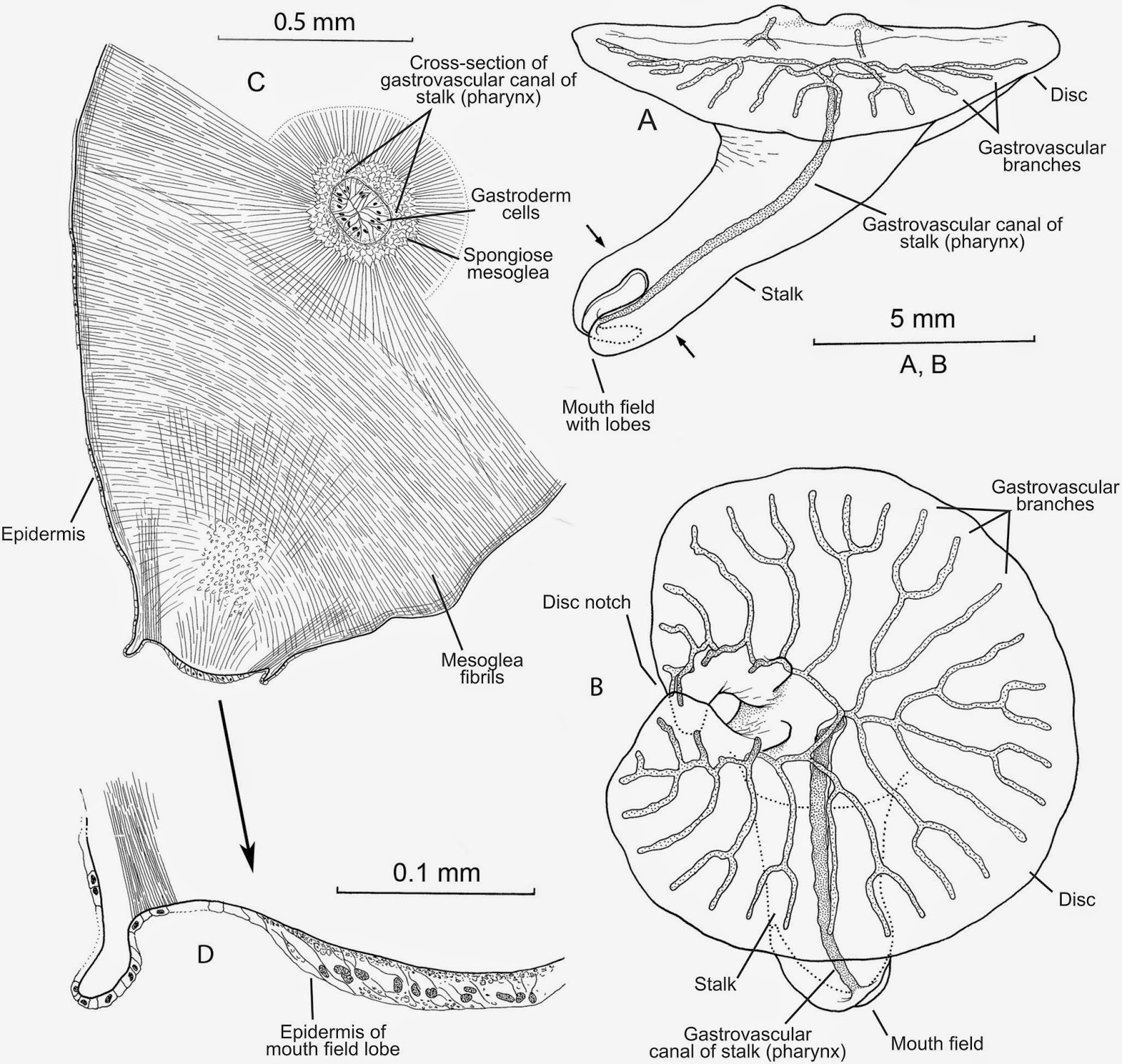
L’arénicole (Arenicola marina) est un ver psammivore, c'est-à-dire qu’il se nourrit de particules accrochées aux grains de sable. Concrètement, il avale plein de sable puis l’estomac fait le tri. Ça leur donne un rôle hyper important puisqu’ils nettoient le sable! Et tout comme les vers de terre, ils laissent des turricules à la surface du sol. Ils aspirent le sable par la bouche ce qui crée une petite dépression à la surface du sol au dessus de leur tête, et de l’autre côté, ils expulsent le sable nettoyé sous forme de tortillon. On peut donc savoir dans quelle direction il est enfoui.
Pas besoin ici d'avoir le ver sous les yeux pour savoir duquel il s’agit !
Dans beaucoup d’environnements, on peut aussi utiliser le critère des traces pour identifier des animaux. Si vous allez régulièrement à la plage, vous avez sûrement dû voir des pêcheurs chercher des vers pour appâter leurs hameçons à l’aide d’une pelle ou d’une bêche : ils s'en vont chercher des vers marins fouisseurs. Ça n’a peut-être pas l’air, mais finalement, c’est très simple d'en trouver quand on sait ce qu’il faut regarder.
L’arénicole (Arenicola marina) est un ver psammivore, c'est-à-dire qu’il se nourrit de particules accrochées aux grains de sable. Concrètement, il avale plein de sable puis l’estomac fait le tri. Ça leur donne un rôle hyper important puisqu’ils nettoient le sable! Et tout comme les vers de terre, ils laissent des turricules à la surface du sol. Ils aspirent le sable par la bouche ce qui crée une petite dépression à la surface du sol au dessus de leur tête, et de l’autre côté, ils expulsent le sable nettoyé sous forme de tortillon. On peut donc savoir dans quelle direction il est enfoui.
Pas besoin ici d'avoir le ver sous les yeux pour savoir duquel il s’agit !
 |
| A gauche : turritule en forme de cœur (St Valentin oblige) laissée par une arénicole (photo prise par nos soins). A droite, schéma de l’arénicole dans son terrier (la tête est à gauche où elle aspire le sable, à gauche, elle expulse), source. |
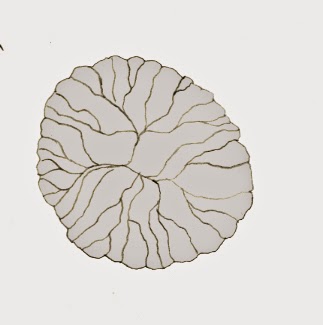
D’autres annélides telles que la célèbre Lanice conchilega (de la famille des Terebellidae parce qu’elles sont très belles) se nourrissent en récoltant les particules fines à l’aide de leurs tentacules. Pour se protéger, elles construisent un tube à partir de débris de coquillages ou de grains de sable agglomérés par du mucus. Seule une petite partie du tube sort du sable et à l’extrémité de celui-ci, des débris de plus petite taille sont utilisés pour construire un panache porte-tentacules. En effet, le ver pose ses tentacules sur chacune de ces extensions qui lui permettent de récupérer la nourriture sur une distance plus longue. Il est assez difficile de sortir l’animal de son tube (il faut un outil pour creuser et être rapide), mais la simple présence de sa construction nous confirme qu’il est bel et bien là.
Puisque des images sont parfois plus parlantes qu’un long discours, voici la partie apparente du tube de la lanice. Si vous y prêtez attention, vous devriez pouvoir en voir assez facilement sur les plages sableuses à marée basse.
Puisque des images sont parfois plus parlantes qu’un long discours, voici la partie apparente du tube de la lanice. Si vous y prêtez attention, vous devriez pouvoir en voir assez facilement sur les plages sableuses à marée basse.
 |
| Tube de Lanice conchilega sur lequel on discerne bien les morceaux de coquillages grossiers pour la partie principale du tube et les morceaux plus fins pour le panache porte-tentacules. Source |
L’ouïe :
Après le sens de la vue, c'est souvent celui de l'ouïe qui est le plus sollicité : en effet, nous l’utilisons pour communiquer dans la vie de tous les jours mais également avec des gens situés à l’autre bout du monde. La nature elle-même n’est pas silencieuse et de nombreux sons peuvent être émis, entendus et décryptés.
Les sons que nous pouvons entendre en nous baladant sont de différents types. Les premiers, sont les sons émis par les animaux pour communiquer entre eux. Il peut s’agir des chants d’alerte « attention, nous ne sommes pas seuls », des chants de parade nuptiale « approche et regarde comme je suis beau gosse », de la communication plus courante entre plusieurs individus (« j’ai faim », « attention intrus »…). Et c’est grâce à ces chants qu’on peut reconnaître les oiseaux de loin, sans avoir besoin de les voir ni de les déranger. Mais il existe aussi les sons liés à l’écologie des bêtes. Un oiseau qui picore dans l’écorce des arbres nous permet, rien qu'à l'oreille, de différencier les espèces. Ainsi, le pic noir et le pic vert ne produisent pas le même son lorsqu'ils martèlent le tronc des arbres pour trouver leur nourriture.
 |
| Sources : là et là |
Mais pensez surtout à un insecte que vous reconnaissez tout de suite au son du vol. Bien sûr ! Le moustique ! Un compagnon indésirable des chaudes nuits d’été. Vous connaissez probablement tellement bien ce bruit que vous n’avez aucun doute sur son émetteur. Bon, mais si le vol est une des manières d’émettre un son, il y a aussi des insectes qui « chantent ». On parle communément de « chant », mais il s'agit en fait de stridulation : un son produit par le frottement de deux surfaces. Par exemple vous saurez tout de suite faire la différence entre un criquet et une cigale rien qu'en les entendant ! D’ailleurs ce sont deux insectes très différents. Si du point de vue évolutif la cigale est proche de la punaise, le criquet, lui, est proche du grillon. Du coup il est assez facile de différencier les deux stridulations, écoutez plutôt :
Grillon : http://iainpetrie.typepad.com/files/grass1.mp3
Criquet : http://www.grammas-tales.com/stuart/cricket2.wav
Chez le grillon ce sont les ailes de la première paire (rappelons que les insectes ont généralement deux paires d’ailes) qui vont se frotter l’une contre l’autre. Alors que chez le criquet, c’est la dernière paire de pattes (celles qui servent à sauter) qui se frotte contre la deuxième paire d’ailes. Deux mécanismes très différents pour des insectes pourtant proches. Il n’est alors pas étonnant que le son produit soit différent. Les experts de ces groupes d’insectes, les orthoptéristes, se basent énormément sur ces « chants » pour distinguer les différentes espèces. On peut d’ailleurs trouver des CD avec les stridulations de différentes espèces pour s’aider et apprendre. Mieux encore, la décomposition de la stridulation des criquets a même déjà été utilisée pour faire des classifications !
Mais il n’y a pas que les criquets et les grillons qui stridulent ! Par exemple le longicorne aussi le fait, mais contrairement à eux, il ne le fait pas pour la drague mais lorsqu’il est inquiet, regardez et surtout écoutez cet exemple (vous pourrez en plus profiter du bourdonnement typique des coléoptères) :
Mais il n’y a pas que les criquets et les grillons qui stridulent ! Par exemple le longicorne aussi le fait, mais contrairement à eux, il ne le fait pas pour la drague mais lorsqu’il est inquiet, regardez et surtout écoutez cet exemple (vous pourrez en plus profiter du bourdonnement typique des coléoptères) :
Cette stridulation est quant à elle provoquée par le frottement des deux premiers segments du thorax (la partie du milieu des insectes qui relie la tête et l’abdomen).
Hormis ces quelques cas, d’autres animaux peuvent être reconnus grâce au son et plus particulièrement grâce aux ultrasons. C’est le cas des chauves-souris. Pour se déplacer, certaines émettent des ultrasons en permanence. Mais pour les entendre, il faut un appareil permettant d’amplifier le son et de le transformer à une fréquence audible pour l'humain. Chaque espèce de chauve-souris émet un son différent, qui permet avec un peu d’expérience de les identifier ! Voici un lien vers le site de VigieNature (dépendant du MNHN) qui explique comment faire, si vous vous sentez l'âme d'un explorateur nocturne.
Hormis ces quelques cas, d’autres animaux peuvent être reconnus grâce au son et plus particulièrement grâce aux ultrasons. C’est le cas des chauves-souris. Pour se déplacer, certaines émettent des ultrasons en permanence. Mais pour les entendre, il faut un appareil permettant d’amplifier le son et de le transformer à une fréquence audible pour l'humain. Chaque espèce de chauve-souris émet un son différent, qui permet avec un peu d’expérience de les identifier ! Voici un lien vers le site de VigieNature (dépendant du MNHN) qui explique comment faire, si vous vous sentez l'âme d'un explorateur nocturne.
Le toucher :
Ce sens est moins utilisé par les naturalistes lors de leurs séances d'identification. Cependant certaines personnes sont plus "tactiles" que d'autres et utilisent beaucoup le toucher pour percevoir leur environnement. Le naturaliste peut aussi utiliser ce sens. Alors pourquoi empêcher les enfants de toucher tous les organismes croisés dans la nature ? Il faudrait presque les y encourager, du moment que cela ne porte préjudice ni à l'organisme étudié, ni à l'observateur !
La parole est à Nicobola ! Anecdote directement sortie des tiroirs pour vous !
Il y a un ver bien connu des étudiants, c’est la nereis. C’est un ver très commun notamment dans la vase et le sable. Elle appartient à la famille des Nereidae. Il existe plusieurs espèces très semblables qui peuvent notamment se différencier grâce au nombre d’yeux, de tentacules et de palpes (des appendices sur la tête). Certains se reconnaissent au comportement comme Platynereis qui vit dans un tube. Une autre méthode pour en identifier certains est la texture. Lors d’un stage d’été, je me rappelle avoir farfouillé dans la vase à la recherche de vers. Le professeur à côté de moi m’indique alors qu’il y a probablement deux espèces : une molle, la nereis commune (Hediste diversicolor) et une autre nereis plus ferme, la Perinereis. Et cette méthode fonctionnait bien. Alors je ne peux pas vous assurer que ça fonctionne seulement sur les Perinereis mais voilà au moins une méthode pour savoir qu’on a bien à faire à deux espèces différentes !
 |
| A gauche une Perinereis (source), à droite, Nereis ou Hediste (source). Ouais, ce sont des vers, et les différencier dans la vase est encore pire… |
En mer aussi on trouve beaucoup d’animaux dont la forme est assez dure à interpréter, ce qui n'aide pas l'identification ! En effet, certains organismes forment plus des masses informes qu’autre chose (c’est surtout l’impression qu’on peut avoir lorsque nous sommes débutant). Ceci arrive plus souvent chez les animaux qui vivent fixés et filtrent les particules en suspension dans l’eau, notamment les éponges (d'autres infos sur les éponges ici et ici) et les ascidies. Cependant ce serait un peu simpliste car les ascidies et les éponges sont des organismes très différents : l’ascidie est proche de nous évolutivement parlant (par rapport, par exemple à l’abeille) alors que les éponges sont probablement les organismes animaux les plus éloignés de nous (enfin, peut-être pas… allez voir ici ! L’éponge est constituée d'un ensemble de canaux et de cavités soutenus dans certains cas par de petites « spicules », une forme de squelette en kit constitué de petites « épines » non attachées entre elles. Tandis que très schématiquement, une ascidie peut être comparée à un sac semi-rigide ponctué de deux trous qui correspondent à des siphons : un pour aspirer l’eau et un autre pour la rejeter (si vous voulez voir ça de plus près, ici, une vidéo faite par nos soins). Elle a aussi une « tunique » assez rigide qui la protège.
Si les deux peuvent grossièrement être confondues à l’œil nu, au toucher la différence devient plus évidente. Premièrement par un toucher superficiel : l’éponge est spongieuse bien sur ! Elle est molle et va se déformer et reprendre doucement sa forme après avoir été pressée. L’ascidie est plus rigide, elle va se déformer mais va donner une impression bien plus ferme et glissante ! Puis elle va très vite reprendre sa forme. Ensuite en pressant plus fort et en dehors de l’eau, l’éponge va rejeter de l’eau partout autour de la zone que l’on presse contrairement à l’ascidie qui ne va la rejeter que par deux orifices, les siphons. Un dernier indice tactilement plus dérangeant, les éponges avec des spicules peuvent être irritantes ! En effet, en les pressant les spicules vont rentrer dans la peau et gratter ! La douleur elle même est donc un moyen d’identifier une éponge !
 |
| A gauche une ascidie coloniale (source), à droite une éponge encroûtante (source)… Le moyen le plus sûr de ne pas confondre est de toucher ! |
En parlant de douleur et d’identification, voici une autre anecdote : il m’est arrivé une fois de chercher des organismes sur des pontons flottants. En effet, le ponton flottant est toujours immergé mais accessible facilement. On y trouve fixés des animaux que l’on ne rencontre habituellement qu’en plongée. Dans une de mes folies aventurières, j’ai voulu y jeter un coup d’œil de très près. Je m’y suis donc rendu à la nage mais malheureusement n’y voyais que des algues (le courant était fort et il était difficile de bien voir ce qu’il y avait). Puis le courant m’a poussé sur le ponton, l’épaule entrant en contact avec ces algues… Premier contact, ça brûle ! Je n’y fais pas trop gaffe… Deuxième contact, ça brûle vraiment ! Eurêka ! Ce n'étaient pas des algues mais des Hydrozoaires ! Parce que si ça brûle c’est que c’est un cnidaire ! La famille des méduses et coraux qui ont des cellules urticantes très spécifiques ! Leur forme « végétale » quant à elle, est plutôt commune aux hydrozoaires ! Je décidais donc d’en prendre sur le bord de plage et cette fois-ci, grâce à ma vue j’ai pu confirmer ma supposition et identifier ça comme un joli Tubularia !
 |
| Dans le remous des vagues dur d’imaginer que ce magnifique animal… est un animal ! (Source) |
A mon tour, à mon tour ! Deux mots avant que Boris ne reprenne la parole après les anecdotes passionnantes de Nico.
Pour continuer dans le domaine du « toucher marin », un petit exemple avec deux petits poissons (oulalalala, j’ai osé prononcer le mot interdit). Il y a quelques années maintenant, j’ai fait un stage sur la faune littorale marine dans lequel j’ai appris énormément sur la biologie, le comportement, ou l’anatomie des bestioles qui composent ces écosystèmes. Comme plusieurs de mes collègues stagiaires, on avait beaucoup de mal à se rappeler la différence entre les blennies et gobies. A part le célèbre argument « parce que je le sais », difficile de mettre des mots sur les différences même si en regardant sur des photos, ce ne sont pas les bêtes les plus mimétiques du monde.
Les confusions étaient surtout due à la biologie de ce type de poisson, les deux espèces étant de petites tailles et se posant préférentiellement sur le fond, dans les recoins ou les flaques. Mais au final, lorsqu’on les touche, pas de doute. Tandis que la blennie est lisse car dépourvue d’écailles (ou écailles rudimentaires), le gobie a des écailles qui lui confèrent un toucher bien plus rugueux. Facile hein ? (Encore faut-il pouvoir les attraper, et alors là, bon courage).
Les confusions étaient surtout due à la biologie de ce type de poisson, les deux espèces étant de petites tailles et se posant préférentiellement sur le fond, dans les recoins ou les flaques. Mais au final, lorsqu’on les touche, pas de doute. Tandis que la blennie est lisse car dépourvue d’écailles (ou écailles rudimentaires), le gobie a des écailles qui lui confèrent un toucher bien plus rugueux. Facile hein ? (Encore faut-il pouvoir les attraper, et alors là, bon courage).
 |
| A gauche : Parablennuis gattorugine (une blennie, photo faite par nos soins), à droite, Gobius paganellus (un gobie, source). |
Place à la botanique ! Au tour de Boris de parler !
On peut également différencier les plantes à l’aide du toucher. Un premier exemple qui me vient à l’esprit et qui vous parlera très facilement, c’est l’ortie (Urtica dioica) qui est bien connue pour les douleurs qu’elle provoque. Cependant, d’autres plantes présentent des caractéristiques moins douloureuses qui peuvent être identifiées rien qu'avec le toucher. Entre autre, la grande consoude (Symphytum officinale) qui possède sur la tige et les feuilles des poils rugueux très facilement identifiables les yeux fermés :
 |
| Source |
D’autres plantes comme celles de la famille des Geraniaceae (les géranium) ou les Lamiaceae (où l’on retrouve la menthe, le romarin…) possèdent des poils plus ou moins duveteux et soyeux. Je me souviens, lors d’un stage, je devais apprendre à différencier le Geranium rotundifolium du Geranium molle au stade de plantule, et pour cela, nous n’avions à notre disposition que les tiges poilues. Il a bien fallut que j’utilise ce que j’avais sous la main pour faire la différence entre les deux ! Car ces deux espèces n’ont pas les mêmes poils sur la tige : le G. molle est plus poilu que le G. rotundifolium.
D’autres plantes, comme les Poaceae, s’identifient assez bien rien qu’en passant la main dessus. Ainsi, la houlque laineuse (Holcus lanatus) et le dactyle aggloméré (Dactylis glomerata) peuvent être confondus lorsqu’ils ne sont pas encore en fleur. Pour les différencier, il suffit de passer les doigts sur la tige : la houlque est beaucoup plus douce au toucher que le dactyle.
D’autres plantes, comme les Poaceae, s’identifient assez bien rien qu’en passant la main dessus. Ainsi, la houlque laineuse (Holcus lanatus) et le dactyle aggloméré (Dactylis glomerata) peuvent être confondus lorsqu’ils ne sont pas encore en fleur. Pour les différencier, il suffit de passer les doigts sur la tige : la houlque est beaucoup plus douce au toucher que le dactyle.
 |
| A gauche le dactyle (source), à droite la houlque (source) |
En hiver, la plupart des arbres de nos régions perdent leurs feuilles (on dit que le feuillage est caduc). Seuls les troncs nus restent accessibles pour le naturaliste… mais tout n’est pas perdu, loin de là ! Il est très facile d’identifier les écorces au toucher. Une écorce lisse sera associée au hêtre (Fagus sylvatica) ou au charme (Carpinus betulus) tandis qu’une écorce rugueuse sera associée au chêne (Quercus robur)… Bien évidement, les informations récoltées à l’aide du sens du toucher sont à mettre en relation avec d’autres informations obtenues à l’aide des autres sens pour une identification complète.
 |
| Écorce rugueuse du chêne pubescent à gauche (source) et écorce lisse du hêtre (source) |
L’odorat :
Après la vue, l'ouïe, le toucher vient l'odorat. Le naturaliste peut être amené à utiliser ce sens bien plus souvent que dans la vie de tous les jours, notamment avec les organismes qu’il peut manipuler. Par exemple beaucoup de plantes et d’animaux rejettent des odeurs particulières pour attirer ou repousser d’autres organismes.
Aurélide a la parole :
Pour reprendre dans les anecdotes littorales, je me souviens d’une sortie à marée basse lors de ce stage sur la faune marine (dont j’ai parlé un peu plus haut). Notre maître de stage nous a appelés, Nico et moi, vers un banc de sable et nous a demandé de sentir, sans se pencher, sans s’approcher du sol, sans creuser, juste là, tous les trois à sniffer l’air. En quelques instants, l’odeur est parvenue à nos narines. Une odeur infecte d’œuf pourri, c’était atroce (et non, ce n’était pas une blague de mauvais goût). Il ne s’agissait pas de matière en décomposition, mais d’un annélide qui porte TRÈS BIEN son nom : Phylo foetida. Un moyen désagréable mais très simple de repérer et identifier la bête.
(D’ailleurs, Nico a fait l’amère expérience de se retrouver avec un de ces vers sous son lit. Les stagiaires avaient trouvé bon de lui faire cette mauvaise blague. Pour le coup, on ne pourra pas mentionner la célèbre phrase « aucun animal n’a été maltraité dans ce tournage », désolés…)
(D’ailleurs, Nico a fait l’amère expérience de se retrouver avec un de ces vers sous son lit. Les stagiaires avaient trouvé bon de lui faire cette mauvaise blague. Pour le coup, on ne pourra pas mentionner la célèbre phrase « aucun animal n’a été maltraité dans ce tournage », désolés…)
Après cette anecdote malodorante, voici un peu de douceur avec Boris :
Evidemment, on peut reconnaître certaines fleurs à leur parfum : le lys (Lilium sp.), la rose (Rosa canina) et tant d’autres que l’on trouve chez le fleuriste. Cependant, les fleurs ne sont pas les seuls organes odorants. Parfois, elles ne sentent rien, ou bien ne sont même pas présentes. Il faut donc chercher d’autres parties de la plante qui possèdent des critères odorants.
C’est le cas par exemple de la mélisse (Melissa officinalis), qui ressemble beaucoup à la menthe odorante (Mentha suaveolens)… tant qu’elle ne porte pas de fleurs. Il suffit de froisser alors les feuilles de la mélisse entre ses doigts pour se rendre compte qu’elle libère un parfum rappelant celui du citron ! Rien à voir avec la menthe…
C’est le cas par exemple de la mélisse (Melissa officinalis), qui ressemble beaucoup à la menthe odorante (Mentha suaveolens)… tant qu’elle ne porte pas de fleurs. Il suffit de froisser alors les feuilles de la mélisse entre ses doigts pour se rendre compte qu’elle libère un parfum rappelant celui du citron ! Rien à voir avec la menthe…
 |
| A gauche la mélisse (source), à droite la menthe (source). |
Le géranium herbe-à-robert (Geranium robertianum) quant à lui est facilement identifiable grâce à l’odeur détestable qu’il répand lorsque ces feuilles sont coupées, ce qui permet de l’identifier facilement parmi d’autres espèces de géranium.
S’il vous arrive de vous promener en bordure de mer, vous avez peut être rencontré cette plante : l’ajonc d’europe (Ulex europaeus), à ne pas confondre avec le genêt à balais (Cytisus scoparius). Bien que ces deux plantes possèdent une morphologie différente, elles ont des fleurs très semblables. Un critère odorant à coup sûr pour trancher est de froisser la fleur entre ses doigts : si elle libère une odeur de noix de coco, c’est que vous avez un ajonc en face de vous !
S’il vous arrive de vous promener en bordure de mer, vous avez peut être rencontré cette plante : l’ajonc d’europe (Ulex europaeus), à ne pas confondre avec le genêt à balais (Cytisus scoparius). Bien que ces deux plantes possèdent une morphologie différente, elles ont des fleurs très semblables. Un critère odorant à coup sûr pour trancher est de froisser la fleur entre ses doigts : si elle libère une odeur de noix de coco, c’est que vous avez un ajonc en face de vous !
 |
| A gauche du genet (source), à droite de l’ajonc (source). |
Une autre plante qui pousse au printemps sur le bord des chemins pourrait passer inaperçue… si elle ne produisait pas une odeur caractéristique dès lors que ses feuilles sont tranchées : il s’agit de l’Alliaire (Allaria petiolata) qui émet une forte odeur d’ail dès que ses feuilles sont coupées. Aucun doute possible alors quant à l’identité de la plante…
Le goût :
Eh oui ! On peut utiliser ce sens pour l'identification des organismes ! Même si on l'utilise tous les jours lorsque l’on mange, il est rarement sollicité pour d'autres raisons. S’il est assez rare d’identifier les animaux par le gout dans un cadre naturaliste (cela signifierait les tuer en général, et tous les animaux identifiables par le gout se retrouvent souvent dans nos assiettes), les plantes et champignons eux peuvent l’être de manière plus aisée ! Et bien qu’un grand nombre d’entre eux soient toxiques pour l’être humain, il n’est pas rare que certains manuels naturalistes conseillent de goûter un petit morceau de l’organisme pour savoir s’il possède un goût particulier (certains champignons non comestibles goûtent fortement le fromage… difficile de se tromper sur l’identification dans ce cas !).
Bien évidement, quand on imagine identifier les plantes au goût, on pense tout de suite aux fruits qu’elles peuvent donner. Mais d’autres parties de la plante peuvent être mastiquées pour aider à l’identification. C’est le cas par exemple du tussilage, ou pas-d’ane (Tussilago farfara) qui peut être difficile à identifier car en été, seules les feuilles subsistent alors que les fleurs sont fanées. En goûtant les feuilles du tussilage, on garde sur la langue un goût de poivre très prononcé.
Une autre plante, Lepidium campestre, possède quant à elle un goût prononcé de… chou-fleur. Même sans les fleurs, il est donc aisé de la reconnaître !
D’autres organismes, qui ne sont pas des plantes mais qui sont aussi des organismes fixés, se reconnaissent souvent à leur goût. Il s’agit des champignons (la plupart des champignons comestibles sont des basidiomycètes). Ainsi, la russule intègre (Russula integra) possède un goût de noisette lorsque sa chair est consommée crue. D’autres champignons peuvent avoir des goûts se rapprochant du poivre, du miel ou encore… du camembert !
Dans le cas des algues, l’espèce Osmundea pinnatifida se reconnait assez facilement par la vue lorsqu’on a un poil d’expérience (par sa couleur, sa forme et aussi l’endroit dans lequel elle se trouve), mais pour l’identifier à coups sûr, il suffit d’en croquer un bout qui a un léger goût d’ail poivré (très bon dans une salade d’ailleurs !).
Les cinq sens nous sont essentiels pour la vie de tous les jours. Si l'un d'entre eux nous fait défaut, il est souvent contrebalancé par le surdéveloppement des autres. Dans le domaine du naturalisme aussi, on retrouve cette complémentarité entre les sens lors de l'identification des organismes, tout comme il existe des cas pour lesquels l'un des cinq sens sera plus utile que les autres. Il y a tout un tas de façons de reconnaître les organismes qui nous entourent. Même si vous n’avez besoin que d’éléments visuels n’hésitez pas à sentir, toucher, écouter, goûter. Cela contribue forcément au processus de mémorisation. D’autant plus que c’est aussi super chouette de pouvoir réunir le plus de critères possibles. Mais aussi, il arrive que la diversité au sein d’un taxon fausse votre identification et vous mette à rude épreuve. Il est alors bien plus sûr de pouvoir combiner le plus d’indices possibles.
Bibliographie :
- Eggenberg S, Möhl A. 2008. Flora vegetativa. Edition Rossolis.
- Knudsen H, Petersen JH. 2005. Les Champignons dans la nature. Edition Delachaux et Niestlé.
- ADER Denis, DUMAS Jacques, HUET Sylvie, in : DORIS, 27/1/2014 : Lanice conchilega (Pallas 1766), http://doris.ffessm.fr/fiche2.asp?fiche_numero=505
- WEBER Matthias, SITTLER Alain-Pierre, REGUIEG Aedwina, CHANET Bruno, in : DORIS, 15/1/2014 : Gobius paganellus Linnaeus, 1758, http://doris.ffessm.fr/fiche2.asp?fiche_numero=1181
La majorité de ces anecdotes proviennent de notre expérience sur le terrain ou de ce que nous avons appris en cours, d’où une biblio courte !
- Eggenberg S, Möhl A. 2008. Flora vegetativa. Edition Rossolis.
- Knudsen H, Petersen JH. 2005. Les Champignons dans la nature. Edition Delachaux et Niestlé.
- ADER Denis, DUMAS Jacques, HUET Sylvie, in : DORIS, 27/1/2014 : Lanice conchilega (Pallas 1766), http://doris.ffessm.fr/fiche2.asp?fiche_numero=505
- WEBER Matthias, SITTLER Alain-Pierre, REGUIEG Aedwina, CHANET Bruno, in : DORIS, 15/1/2014 : Gobius paganellus Linnaeus, 1758, http://doris.ffessm.fr/fiche2.asp?fiche_numero=1181
La majorité de ces anecdotes proviennent de notre expérience sur le terrain ou de ce que nous avons appris en cours, d’où une biblio courte !





































































.PNG)

































%2C%2BGreater%2BFlamingo%2B%2B(2).jpg)